Plankton dynamics
Research on the lakes of the McMurdo Dry Valleys (MCM) began with the advent of the International Geophysical Year (IGY) in the late 1950’s. IGY research revealed the physical/chemical nature of the lakes, showing that they were the only year-round liquid water environments on the continent, and inferred that the biological systems in the permanently ice-covered lakes must possess novel physiological strategies that allow them to survive at low temperature and under extended darkness. The seminal studies during IGY provided the framework for subsequent hypotheses driven research which now forms the basis for the ongoing MCM LTER lake program (now in its 15th season; www.mcmlter.org) and the MCM dry valley lake Microbial Observatory (http://www.homepage.montana.edu/~lkbonney/).
Data collected on Lakes Bonney, Hoare and Fryxell (all in the Taylor Valley) as part of the MCM LTER have shown that most organisms in the lakes are not just "surviving the extremes" but are actively feeding, growing and reproducing. As such, they are ecosystems in which we can identify and begin to understand physiological and genomic adaptations in the context of one of the most extreme environments on our planet. Unfortunately, almost all "hands-on" research on the MCM lakes has been restricted to the austral spring and early summer (~October through January) when logistical support has allowed access to the area.
Although studies during the spring/ summer period have yielded a quantum increase in our understanding of the lakes, the unique aspects of physiological adaptation, biodiversity and ecosystem function during the permanently cold and prolonged darkness of the Antarctic winter will never be understood without extended season research. Through an International Polar Year (IPY) grant from NSF-OPP we were able to conduct the first research on these lakes during the transition from 24 hour sunlight to total darkness between November 2007 and April 2008.
The overarching theme of this research was that the onset of darkness induces a cascade of physiological changes that alters the functional roles of autotrophic and heterotrophic microplankton within the lakes. This overarching theme was addressed through an interdisciplinary and international study of selected biological components of the lakes using genomic and physiological tools to understand not only how individual organisms survive, but how they control ecosystem function during this seasonal transition. This theme is directly relevant to MCM LTER and IPY objectives, and the data will be critical to our knowledge of how polar organisms survive and function within high latitude ecosystems characterized by extreme cold and prolonged darkness. The theme also contributes directly to the international Scientific Committee on Antarctic Research (SCAR) program "Evolution and Biodiversity in the Antarctic" (EBA; http://www.eba.aq/).
By virtue of perennial ice covers, low advective stream flow, and saline bottom waters, lakes in the MCM are permanently stratified and have a unique and diverse set of thermodynamic and geochemical conditions. Water temperatures range from -5 to 6° C and fluctuate less than 26° C seasonally. Superimposed on these stable cold temperatures is a low light environment where measurable photosynthetically available radiation (PAR) penetrates the ice only from early October to mid-March.
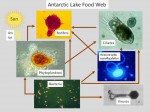
Data collected during spring and early summer as part of the MCM LTER and Microbial Observatory programs have revealed multiple, well-developed phytoplankton and bacterial maxima associated with nutrient gradients down the water column. Strong microbial linkages have been defined in all of the MCM lakes and point to the tight coupling of carbon and other nutrients and the reliance of the system on phytoplankton photosynthesis during the short summer period (Figure 1).
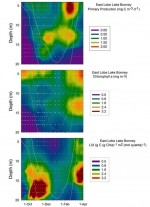
We currently know little about foodweb dynamics in late summer, autumn, or the long dark winter. Based on the summer to winter transition data processed thus far, we can now produce a picture of annual bacterial and phytoplankton productivity in all lakes. Preliminary analyses of these data show that chlorophyll-a continues to increase after phototrophic primary production has ceased (Figure 2). This is due in part to cryptomonad phytoplankton switching from light driven metabolism to phagotrophy on bacteria, which is a winter survival mechanism shown to occur in other Antarctic lakes. A mid-season drop in photosynthetic efficiency apparently occurs as the result of phosphorus deficiency by the phytoplankton, a fact we corroborated through nutrient bioassay experiments which showed significant stimulation of light mediated 14CO2 incorporation by the addition of phosphorus. A late season increase in photosynthetic efficiency apparently results from the leaching of phosphorus from sediment released from the ice permanent cover.
Clearly, IPY-funded research into the polar night has provided the MCM LTER with a window of opportunity to collect new data during a time of year that that will allow us to refine our understanding of lake ecosystem processes. The permanently icecovered lakes being studied by the MCM LTER are the only known environments on the Antarctic continent that contain liquid water throughout the year. As such, these lakes are truly an oasis for life in what would otherwise appear to be an inhospitable environment.
John C. Priscu, Department of Land Resources and Environmental Sciences, Montana State University, Bozeman, MT
 Enlarge this image
Enlarge this image